- Research Article
- Open access
- Published:
Influence of bimanual exercise on muscle activation in post-stroke patients
ROBOMECH Journal volume 6, Article number: 14 (2019)
Abstract
Brain damage due to stroke often leaves survivors with lateral functional motor deficits. Bimanual rehabilitation of the paretic arm is an active field of research aimed at restoring normal functionality by making use of the complex neural bindings that exist between the arms. In search of an effective rehabilitation method, we introduced a group of post-stroke rehabilitation patients to a set of bimanual motion tasks with inter-manual coupling and phasing. The surface EMG profiles of the patients were compared in order to understand the effect of the motion conditions. The paretic arms of the patients were more strongly affected by the task conditions compared with the non-paretic arms. These results suggest that in-phase motion may activate neural circuits that trigger recovery. Coupling also had an effect on behavior, but the response of patients was divided between those whom coupling helped or hindered.
Introduction
Stroke survivors are often left with functional impairment of the arm contralateral to the brain lesion. Accordingly, in daily tasks that they previously performed using both hands, they tend to use only the arm that is less affected by stroke [1]. Bimanual training has been proposed as a therapy for rehabilitating the motor function of the paretic arm [2,3,4] with the aim of restoring normal quality of life. Constraint-induced motion therapy [5], where the use of less-affected arm is restrained, has been clinically tested [6] and the effect has also been identified with brain imaging [7, 8], but, nevertheless, the method suffers from the need to deploy a therapist, just like conventional therapy. Also, isometric tasks with bimanual force-control models, sometimes combined with with brain stimulation, have been proposed [9,10,11], however, the actual rehabilitation applications are yet to be introduced.
Recovery after stroke is connected to the reorganization of surviving neural structures [12, 13]. Bimanual training seeks to employ many forms of bilateral neural coupling mechanisms to enhance and harness the plasticity of the central nervous system [14, 15]. However, to date, few detailed analyses of bimanual motion training have been reported, even though the effectiveness of bimanual training for recovery from motor paralysis is widely recognized [16, 17]. In order to develop therapy methods, the underlying mechanisms of bimanual mechanisms need to be investigated further [18, 19].
Current bimanual therapy relies heavily on specific training devices [20, 21]. These devices are based on passive mechanics, in which the system is driven by forces generated by the patient [22,23,24] or by active robotics [25, 26] to provide symmetric repetitive motion patterns for arm movement. These devices are, however, unsuitable for investigating the biological mechanisms underlying bimanual motion training because they can only provide simple bimanual motions [27].
Both computation and theoretical approaches are applied to deeper understanding on the mechanism of recovery and the role of bimanual motions. Cheung et. al. discussed the changes of muscle synergy after stroke comparing the muscle synergies on paretic and non-paretic arms [28, 29]. They showed that merging and fractionation of the spatial factors of the muscle synergy occurred depending on the recovery stage after stroke. Kantak et. al., proposed a theory-guided taxonomy of bimanual actions for quantifying the coordination of bimanual motion depending on the task in the real-world [30]. Instead of these discussions, the changes of muscle activities during different types of bimanual motions are still unclear.
This study examines the dependence of patients’ muscle activities on the type of bimanual motion using a system that allows for several types of bimanual motion for hemiplegia patients. It is known that the motion phase between paretic and non-paretic arms and paretic motion support by the non-paretic arm are critical factors in bimanual training [31,32,33,34,35]. We used the dual-steering rehabilitation system (DsRS) to create several types of bimanual motions. The DsRS consists of two wheels and a mechanical linkage that can change the connection between the two wheels among three training states: in-phase coupling, anti-phase coupling, and independent motion. When the wheels are mechanically coupled, the non-paretic arm motion can assist the motion of the paretic arm. The mechanism is described in detail below.
We can quantify differences between the influences of the bimanual motion types based on their associated muscle activities as measured by electromyography (EMG) signals. EMG signals are now frequently used as a key physiological marker for observing the features of motion paralysis in post-stroke patients [29, 36,37,38].
In this study, we will quantify the immediate changes of EMG signals depending on the difference of bimanual motion type. Post-stroke patients were asked to perform four types of bimanual motions across two phase and two coupling states using DsRS. Phase, which refers to the movement of the arms with respect to each other, is physiologically important. In-phase movement occurs when homologous muscles work simultaneously in both limbs [39]. Additionally, humans have a natural preference for in-phase movements over anti-phase movements [31, 32]; as the frequency of motion increases, humans tend to switch from anti-phase to in-phase motion. However, this preference is not exclusive because anti-phased motion patterns are a natural part of locomotion [33]. Mechanical coupling of the arms is another common feature in many bimanual training devices [34]. Mechanical coupling distributes the load between the arms, while loads would otherwise be divided between each arm independently. Furthermore, proprioceptive limitations associated with the stroke-affected arm suppress its motor function [35]. Mechanical coupling [23] may increase the sensory information required to control the paretic arm [14].
The remainder of this paper is organized as follows. In “Experimental setup” section, we describe the details of the DsRS mechanism and the types of bimanual motion that the DsRS can provide. We then describe the state of the subjects who participated in the study. In “Experimental protocol” section , we introduce two indices to quantify differences in EMG data dependent on the bimanual motion type. In “Experimental results” section , we go on to discuss the experimental results, showing that EMG patterns of paretic arms depend on the type of the bimanual motion patterns, though the EMG patterns of non-paretic arms remain relatively uniform across those same tasks. Lastly, in “Discussion” section we discuss the bimanual training model.
Experimental setup
In this section, we introduce the device used to create the aforementioned types of bimanual motions. We also characterize the state of the post-stroke patients who participated in the study.
Dual-steering rehabilitation system designed to create various types of bimanual motions
Patients with hemiplegia were asked to perform various types of bimanual motions using the dual-steering rehabilitation system (DsRS) shown in Fig. 1a. This device consists of two steering wheels and a mechanical clutch that can change the state of the mechanical connection between the two wheels to one of three modes, in-phase, anti-phase, and independent modes, as shown in Fig. 1b. While in the in-phase mode, the two wheels are mechanically connected, and when one is rotated by hand, the other rotates in the same direction. In contrast, while in the anti-phase mode, the second wheel rotates in the opposite direction when the first wheel is rotated by hand. While in the independent mode, the two wheels are free to rotate independently. This mechanism is useful for creating different phases of bimanual motions that enable paretic motions to be supported by the non-paretic arm. The rotation of the wheels is measured by encoders.

a Dual-steering rehabilitation system, the wheels of which can be mechanically uncoupled or coupled. b The movement patterns performed in the experiments: in-phase, where hands move in opposite directions; and anti-phase, where hands move in the same direction
Subjects
Eleven post-stroke rehabilitation inpatients (age, M = 74.7, SD = 9.3 years; sex, M = 7, F = 4) were recruited to volunteer for this study. All of the patients had been diagnosed with stroke (see details in Table 1). EMG recordings were made as the patients used the device over a mean of 76 days (SD = 41) post-stroke. The inclusion criteria were the ability to grip and rotate a steering wheel \(180^{\circ }\) independently with each of their arms. The patients were assessed using the Stroke Impairment Assessment Set (SIAS) [40] two times in total, once before and once after the experimental sessions. The SIAS scores of patient motor function performance on a scale from 0 to 5 were scored separately for proximal and distal function, which are measured by their reaching ability and voluntary finger motion, respectively.
All experimental sessions were conducted at the National Center for Geriatrics and Gerontology (NCGG) in Aichi Prefecture, Japan. All experimental protocols were approved by the ethics committee of NCGG.
Experimental protocol
In this section, we introduce the experimental protocol for recording EMG data during DsRS training and introduce two indices to analyze the immediate changes in EMG among types of bimanual motion. One of the indices is used to compute the variance of EMG recordings among the different types of bimanual motions. The other is an index for computing similarities between the different EMG sets.
Experimental protocol
Patients were asked to perform the four types of bimanual motions listed in Fig. 2a combining phase and coupling states. The experimental sessions followed a block design. As described in Fig. 2b, each task block of 20 s was separated by 40 s of rest to avoid physical fatigue. Each session consisted of four tasks, which were repeated three times in a pseudo-randomized order. Prior to the experiments, subjects did light elbow flexing exercises as a warm-up and some rotations on the wheels to familiarize themselves with the device.

a The four task conditions defined as combinations of the phase and coupling conditions. For a given task, the same movement is repeated five times. b In the block design of the experiment, each of the four tasks was repeated five times in a pseudo-random order
Method for quantifying EMG differences among bimanual motions
EMG data recorded during the experiments were analyzed according to the following protocol:
-
1.
Activities of the nine muscles in each side described in Fig. 3a were monitored in accordance with the guidelines of the Surface Electromyography for the Non-Invasive Assessment of Muscles Project [41].
-
2.
The data that contributed less to creating the motions were removed [42].
-
3.
The profiles were extracted for the selected muscles to represent the muscle activation in motion patterns.
-
4.
The variance of the EMG profiles among the different types of bimanual motion were computed from the data.
-
5.
The similarity between paretic and non-paretic arm EMG data for lateral activation similarity (LAS) was computed, and muscle activation similarities between different coupling states (\(\hbox {MAS}_C\)) and between the different phases (\(\hbox {MAS}_P\)) were also computed.
Fig. 3 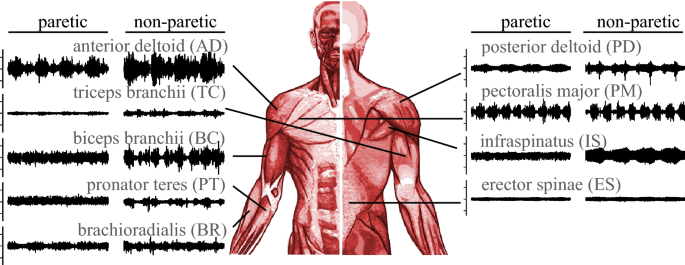
Placement of the EMG sensors and corresponding EMG signals during four cycles of in-phase motion. The EMG signal duration is 20 s and one tick in amplitude is 20 mV

In the experiments, we monitored the activity of nine muscles, as described in Fig. 3. Figure 3 shows an example of control muscle activities during DsRS use. In the case of this particular subject and task, the anterior deltoid was activated in response to DsRS control motion while the triceps was poorly activated. This result implies that the nine measured muscles are not always activated during DsRS use. In trials such as these, incorrect results can be derived from similarity and variance calculations if we use the EMG data based on muscles that did not contribute to the assayed motions [42]. The muscles activated by this motion depend mainly on the individuals assessed in our experiments. Therefore, we removed data on muscle activation for individuals in the first process of analysis using the method proposed by Costa et al. [42]. A flowchart describing this method is provided in Fig. 4b. In this method, the contribution of each muscle is quantified using the EMG amplitude and frequency.

a Dataflow explaining how the measurements from subjects are characterized as analysis parameters. b Detailed explanation of muscle selection process
The extraction of the surface EMG profiles started with preprocessing using a high-pass filter (4th-order Butterworth at 20 Hz) to remove motion artifacts. The filtered signals were rectified by taking absolute values of each datapoint and smoothing the input with a low-pass filter (4th-order Butterworth at 5 Hz). We shifted the half phase of motion in anti-phase motions to align the data according to the same motions. The profiles were obtained by slicing the signals according to changes in the direction of rotation. Slices belonging to the same task within a session were averaged for each muscle.
Description of EMG data
After obtaining EMG profiles, we computed the variances and similarities among the four different types of bimanual motion patterns described in Fig. 2a. To compute these values, we describe the EMG data in the following way:
Here, \({}^*{M}_+\) is the matrix of the data for the selected EMG signal. \(*\) can be P or N, which indicate the paretic or non-paretic arm data, respectively. \(+\) can be IC, IU, AC, or AU, which indcate the type of bimanual motions (See Fig. 2a). For example, \(^P{M}_{IC}\) represents the EMG data set for the paretic arm during in-phase coupled motion, and \(^N{M}_{AU}\) represents the EMG data set for the non-paretic arm during anti-phase uncoupled motion. \(^*{m}_{i_+}\) represents the vector of a selected EMG dataset (i.e., one of the measured nine muscle EMG signals). l represents the number of selected muscles. \(^*sm_{ij_+}\) represents the elements of the vector. k represents the number of data points in the sampling data. We use the same muscles on both arms that contribute to both paretic and non-paretic arm motions.
Quantification of muscle activity variability between the different types of motions
First, we analyzed the EMG data to assess the variability in muscle activation between the paretic and non-paretic arms during the four modes of bimanual motion tasks. Figure 5a describes a prominent example of the difference in muscle activation between the paretic and non-paretic arms. The anterior deltoid muscle of the non-paretic arm can be seen to be activated in almost the same way across the four different modes of bimanual tasks, while the muscle activation of the paretic arm differed among the four modes, especially between the in-phase and anti-phase motions. This result suggests that the muscle activity pattern changes across different motions.

a Example EMG profiles of the anterior deltoid muscles in different tasks during upwards motion. These profiles show that the anterior deltoid muscle of the non-paretic arm was activated in almost the same way across the four different types of bimanual motion, while the corresponding muscle of the paretic arm was activated in different ways depending on the type of bimanual motion. b Statistical analysis of the variability of muscle activities on the non-paretic and paretic arms across all patients’ trials. There was significantly higher dispersion for muscle activation of the paretic arms. The bars represent means, with whiskers representing \(\pm \hbox {SD}\)
To quantify the variability of the muscle activities among the different types of bimanual motion, we used the following equations:
where \(*\) can be P or N, as described for the data matrix above.
Muscle activation similarity
Another index we use in this paper is activation similarity (AS). AS is used to compare the similarities between the two data matrixes represented by \(^*{M}_+\) in (1). For example, the similarity between the paretic and non-paretic arms for in-phase coupled motion can be quantified as follows:
where \(r(*^1,*^2)\) is the Pearson’s coefficient of correlation between the vectors \(*^1\) and \(*^2\) and \(\overline{^{{P}}{m_{_{IC}}}}\) and \(\overline{^{{N}}{m_{_{IC}}}}\) describe the averages of the elements of the vectors \(^{{P}}{{m}_{j_{{IC}}}}\, {\text{and}}\, ^{{N}}{{m}_{j_{{IC}}}}\), respectively.
The muscle activation similarity AS between the paretic and non-paretic arms under the same condition can be used to assess how much the motion of the non-paretic arm is controlled in a healthy way throughout that motion. In contrast, AS for the paretic arm between the different conditions can quantify muscle activation differences among different conditions. In this research, we define AS between the paretic and non-paretic arms in the coupled condition as lateral activation similarity (LAS), while that between the different conditions is defined as muscle activation similarity (MAS).
Experimental results
In this section, we analyze the experimental results with seven patients using the indices introduced in the previous section. We show that the variance of the EMG patterns for the paretic arm among the different types of bimanual motions are larger than those for the non-paretic arm. Furthermore, EMG patterns during coupled in-phase bimanual motion exhibit higher similarity with the EMG pattern of the non-paretic arm.
EMG pattern variance
As described in Fig. 5a in the previous section, the muscle activation patterns differed among the bimanual motion conditions. We quantify the variability in muscle activation pattern using the index \(^{*}\mathrm{D}\) introduced in "Experimental protocol" section. Figure 5b summarizes the statistical analysis for variability of muscle activation. To compute the variability, we applied the mean of Eq. 2 on the selected muscles that contributed to generating the motions, as discussed in the Experimental protocol section; the selected muscles for each patient are summarized in Fig. 6. The variability in muscle activities for the paretic arm was significantly higher than that for the non-paretic arm, suggesting that muscle activation of the paretic arm differs considerably across the bimanual motion types.

Probability of each patient’s muscles being used to represent a task according to the muscle selection method [42]
Muscle activities of the paretic arm depend on the type of bimanual motion
Another research objective is to determine which bimanual motions create the most appropriate EMG patterns with respect to patterns distributed among the different types of bimanual motions. We assessed this objective using the index described by Eq. (5) to determine which bimanual motion type among the four is most suitable for bimanual training.
LAS can quantify the degree of similarity in muscle activations between the paretic and non-paretic arms during the tasks. Figure 7 shows LAS according to phase condition in the experiments. LAS takes significantly lower values when measured in anti-phase compared with those measured during in-phase motion. LAS, however, fails to characterize higher similarity in either coupling condition, as illustrated in Fig. 8.

Lateral activation similarity (LAS) indicates that the contralateral similarity in EMG signals measured for anti-phase motion is lower than that measured for in-phase motion in both coupled and uncoupled tasks. The whiskers represent \(\pm \hbox {SD}\)

Lateral activation similarity (LAS) indicates that the contralateral similarity in EMG measured under coupling is from the same as that measured under uncoupled tasks. The whiskers represent \(\pm \hbox {SD}\)
MAS is a unilateral comparison of muscle activations according to coupling condition. Figure 9 illustrates that the similarity of activations in the paretic arm is reduced under coupling. This result, together with the previous result Fig. 8, indicates that coupling affects the paretic side, but neither the coupled nor uncoupled condition does not systematically increase the similarity. Instead, there is variation between trials; for some trials, coupling hinders similarity, while for others it enhances the similarity in the activation patterns.

Muscle activation similarity (MAS), a parameter comparing EMG similarity between coupled and uncoupled conditions, indicates lower similarity among tasks for the paretic side. The whiskers represent \(\pm \hbox {SD}\)
Discussion
Bimanual training is a widely practiced method to promote recovery from post-stroke motion paralysis, but the mechanism of recovery remain poorly understood. To identify the mechanism behind motion recovery through bimanual training, we introduced two indices to analyze EMG signals associated with four types of bimanual motion.
\(^{*}\mathrm{D}\) in Eq. (2) represents the dispersion of the EMG pattern among different types of bimanual motions. The experimental results described in Fig. 5 show that muscle activation patterns differed among the types of bimanual motion. This result implies that the muscle activation control is tuned automatically, even though the same motions were performed. This muscle activation tuning may be responsible for triggering recovery from motion paralysis.
The analysis using index AS in Eq. (5) suggests the preferential bimanual motion creates similar EMG patterns between paretic and non-paretic arms. Figures 5 and 7 suggest that for in-phase motions, muscle activations are closer between the paretic and non-paretic arms.
One of the important reasons why muscle activities of the paretic arm differ according to bimanual motion type could be the signal processing that occurs in biological control systems. This type of signal processing can be modeled like the neuro-synergy system described in Fig. 10 [43, 44], where information acquired from the environment is gradually symbolized to reduce its dimensions, while control signals are created from this symbolized information to create behaviors. The notions of sensor synergy [45,46,47] and muscle synergy [48,49,50,51,52] represent the input and output processes of the neuro-synergy system, respectively.
We can consider that a stroke damages the middle of this signal processing flow, and the signal accordingly becomes stuck at some intermediate point, thereby causing motion paralysis. The results of this study suggest that appropriate bimanual motion can stimulate improved muscle synergy by circumventing the damaged or damage-affected circuitry.
These results also suggest the importance of careful observation in creating a bimanual rehabilitation exercise menu best suited to each patient’s condition. Statistical analysis suggests that in-phase motion can enhance the appropriate muscle activity changes for the majority of patients. However, there were patients for which anti-phase motions better elicited the appropriate muscle activities. Rehabilitation must be designed to meet each patient individually.
Conclusion
In this study, we developed a system that encourages four types of bimanual motions for post-stroke hemiplegia patients in order to clarify the mechanism behind effective recovery through bimanual motion training. We measured EMG signals and quantified the dispersion of EMG patterns among different bimanual motion patterns and similarities in EMG patterns between the paretic and non-paretic arms for the same motions.
The results described in Fig. 5 show that the different types of bimanual motions create different EMG patterns for the paretic arm, while almost the same EMG patterns appear for the non-paretic arm. Furthermore, Fig. 7 shows that in-phase coupled motion promotes the most appropriate EMG pattern for the majority of patients. However, the results also suggest that the bimanual motion pattern that promotes the most appropriate EMG pattern is patient-dependent, implying that the best training motion for each patient should be carefully determined.
These results stimulate our interests on the difference of muscle activities between bimanual and unilateral motions, although we focused on the muscle activities of bimanual motions during early rehabilitation stage. The role of unilateral motion may grow during later stages of rehabilitation when aiming for “motion separation”, independence of motions between each arms.
For expanding the scope of rehabilitation, inclusion of patients with more sever hemiparesis is also important, e.g. by means of helping the paretic hand to grasp the device [53]. The experimental results also encourage for further investigation on how bimanual motion could be applied to invigorate the creation of the muscle activities on a completely paralyzed arm.
The biological control model described in Fig. 10 is useful for understanding the state of post-stroke neural systems. We are now in discussions to create a rehabilitation system that uses this model to identify the most appropriate patient-specific bimanual training motions thereby encouraging recovery by properly monitoring EMG changes throughout bimanual motion training.

The neuro-synergy system represents signal processing in a biological control system. In biological control systems, information acquired from the environment is gradually symbolized to reduce its dimensions, while control signals are created from this symbolized information to create behaviors. Motion paralysis is caused by weakened or diminished control signals. The results suggest that appropriate bimanual motion can help activate local control loops, resulting in proper muscle activation
Availability of data and materials
Please contact author for data requests.
References
Bailey RR, Klaesner JW, Lang CE (2015) Quantifying real-world upper-limb activity in nondisabled adults and adults with chronic stroke. Neurorehabilit Neural Repair 29(10):969–978
Latimer CP, Keeling J, Lin B, Henderson M, Hale LA (2010) The impact of bilateral therapy on upper limb function after chronic stroke: a systematic review. Disabil Rehabil 32(15):1221–1231
Van Delden A, Peper C, Beek PJ, Kwakkel G (2012) Unilateral versus bilateral upper limb exercise therapy after stroke: a systematic review. J Rehabil Med 44(2):106–117
Hatem SM, Saussez G, della Faille M, Prist V, Zhang X, Dispa D, Bleyenheuft Y (2016) Rehabilitation of motor function after stroke: a multiple systematic review focused on techniques to stimulate upper extremity recovery. Front Hum Neurosci 10:442
Taub E, Uswatte G, Pidikiti R et al (1999) Constraint-induced movement therapy: a new family of techniques with broad application to physical rehabilitation—a clinical review. J Rehabil Res Dev 36(3):237–251
Winstein CJ, Miller JP, Blanton S, Taub E, Uswatte G, Morris D, Nichols D, Wolf S (2003) Methods for a multisite randomized trial to investigate the effect of constraint-induced movement therapy in improving upper extremity function among adults recovering from a cerebrovascular stroke. Neurorehabil Neural Repair 17(3):137–152
Wittenberg GF, Schaechter JD (2009) The neural basis of constraint-induced movement therapy. Curr Opin Neurol 22(6):582–588
Sheng B, Lin M (2009) A longitudinal study of functional magnetic resonance imaging in upper-limb hemiplegia after stroke treated with constraint-induced movement therapy. Brain Inj 23(1):65–70
Kang N, Cauraugh JH (2017) Bilateral synergy as an index of force coordination in chronic stroke. Exp Brain Res 235(5):1501–1509
Patel P, Lodha N (2019) Dynamic bimanual force control in chronic stroke: contribution of non-paretic and paretic hands. Exp Brain Res 237(8):2123–2133
Jin Y, Lee J, Oh S, Celeste Flores Gimenez M, Yoon B (2019) Noninvasive brain stimulation over the m1 enhances bimanual force control ability: a randomized double-blind sham-controlled study. J Motor Behav 51(5):521–531
Grefkes C, Ward NS (2014) Cortical reorganization after stroke: how much and how functional? Neuroscientist 20(1):56–70
Kwakkel G, Veerbeek JM, van Wegen EE, Wolf SL (2015) Constraint-induced movement therapy after stroke. Lancet Neurol 14(2):224–234
Cauraugh JH, Summers JJ (2005) Neural plasticity and bilateral movements: a rehabilitation approach for chronic stroke. Prog Neurobiol 75(5):309–320
Sainburg R, Good D, Przybyla A (2013) Bilateral synergy: a framework for post-stroke rehabilitation. J Neurol Transl Neurosci 1(3):1025
Meng G, Meng X, Tan Y, Yu J, Jin A, Zhao Y, Liu X (2018) Short-term efficacy of hand-arm bimanual intensive training on upper arm function in acute stroke patients: a randomized controlled trial. Front Neurol 8:726
Chen P-M, Kwong PW, Lai CK, Ng SS (2019) Comparison of bilateral and unilateral upper limb training in people with stroke: a systematic review and meta-analysis. PloS one 14(5):0216357
Sheng B, Zhang Y, Meng W, Deng C, Xie S (2016) Bilateral robots for upper-limb stroke rehabilitation: state of the art and future prospects. Med Eng Phys 38(7):587–606
Winstein C, Varghese R (2018) Been there, done that, so what?s next for arm and hand rehabilitation in stroke? NeuroRehabilitation 43(1):3–18
Van Delden AL, Peper CL, Kwakkel G, Beek PJ (2012) A systematic review of bilateral upper limb training devices for poststroke rehabilitation. Stroke Res Treat 2012:972069. https://doi.org/10.1155/2012/972069
Brackenridge J, Bradnam LV, Lennon S, Costi J, Hobbs DA (2016) A review of rehabilitation devices to promote upper limb function following stroke. Neurosci Biomed Eng 4(1):25–42
Whitall J, Waller SM, Silver KH, Macko RF (2000) Repetitive bilateral arm training with rhythmic auditory cueing improves motor function in chronic hemiparetic stroke. Stroke 31(10):2390–2395
Raghavan P, Aluru V, Milani S, Thai P, Geller D, Bilaloglu S, Lu Y, Weisz DJ (2017) Coupled bimanual training using a non-powered device for individuals with severe hemiparesis: a pilot study. Int J Phys Med Rehabil 5(3):404
Treadway E, Gan Z, Remy CD, Gillespie RB (2018) Toward controllable hydraulic coupling of joints in a wearable robot. IEEE Trans Robot 34(3):748–763
Simkins M, Byl N, Kim H, Abrams G, Rosen J (2016) Upper limb bilateral symmetric training with robotic assistance and clinical outcomes for stroke: a pilot study. Int J Intell Comput Cybern 9(1):83–104
Hesse S, Schulte-Tigges G, Konrad M, Bardeleben A, Werner C (2003) Robot-assisted arm trainer for the passive and active practice of bilateral forearm and wrist movements in hemiparetic subjects1. Arch Phys Med Rehabil 84(6):915–920
Gassert R, Dietz V (2018) Rehabilitation robots for the treatment of sensorimotor deficits: a neurophysiological perspective. J Neuroeng Rehabil 15(1):46
Cheung VC, Piron L, Agostini M, Silvoni S, Turolla A, Bizzi E (2009) Stability of muscle synergies for voluntary actions after cortical stroke in humans. Proc Natl Acad Sci 106(46):19563–19568
Cheung VC, Turolla A, Agostini M, Silvoni S, Bennis C, Kasi P, Paganoni S, Bonato P, Bizzi E (2012) Muscle synergy patterns as physiological markers of motor cortical damage. Proc Natl Acad Sci 109(36):14652–14656
Kantak S, Jax S, Wittenberg G (2017) Bimanual coordination: a missing piece of arm rehabilitation after stroke. Restor Neurol Neurosci 35(4):347–364
Kelso J (1984) Phase transitions and critical behavior in human bimanual coordination. Am J Physiol Regul Integr Comp Physiol 246(6):1000–1004
Mechsner F, Kerzel D, Knoblich G, Prinz W (2001) Perceptual basis of bimanual coordination. Nature 414(6859):69
Patrick SK, Noah JA, Yang JF (2009) Interlimb coordination in human crawling reveals similarities in development and neural control with quadrupeds. J Neurophysiol 101(2):603–613
Sleimen-Malkoun R, Temprado J-J, Thefenne L, Berton E (2011) Bimanual training in stroke: how do coupling and symmetry-breaking matter? BMC Neurol 11(1):11
Torre K, Hammami N, Metrot J, van Dokkum L, Coroian F, Mottet D, Amri M, Laffont I (2013) Somatosensory-related limitations for bimanual coordination after stroke. Neurorehabil Neural Repair 27(6):507–515
Clark DJ, Ting LH, Zajac FE, Neptune RR, Kautz SA (2009) Merging of healthy motor modules predicts reduced locomotor performance and muscle coordination complexity post-stroke. J Neurophysiol 103(2):844–857
Safavynia S, Torres-Oviedo G, Ting L (2011) Muscle synergies: implications for clinical evaluation and rehabilitation of movement. Top Spinal Cord Inj Rehabil 17(1):16–24
Roh J, Rymer WZ, Perreault EJ, Yoo SB, Beer RF (2012) Alterations in upper limb muscle synergy structure in chronic stroke survivors. Am J Physiol Heart Circ Physiol 109(3):768–781
Cohen L (1971) Synchronous bimanual movements performed by homologous and non-homologous muscles. Percept Motor Skills 32(2):639–644
Chino N, Sonoda S, Domen K, Saitoh E, Kimura A (1994) Stroke impairment assessment set (sias). Jpn J Rehabil Med 31(2):119–125
Hermens HJ, Freriks B, Merletti R, Stegeman D, Blok J, Rau G, Disselhorst-Klug C, Hägg G (1999) European recommendations for surface electromyography. Roessingh Res Dev 8(2):13–54
Costa A, Itkonen M, Yamasaki H, Alnajjar F, Shimoda S (2017) Importance of muscle selection for EMG signal analysis during upper limb rehabilitation of stroke patients. In: 2017 39th Annual international conference of the IEEE engineering in medicine and biology society (EMBC), pp 2510–2513. https://doi.org/10.1109/EMBC.2017.8037367
Csete M, Doyle J (2004) Bow ties, metabolism and disease. Trends Biotechnol 22(9):446–450
Okajima S, Tournier M, Alnajjar FS, Hayashibe M, Hasegawa Y, Shimoda S (2018) Generation of human-like movement from symbolized information. Front Neurorobot 12:43. https://doi.org/10.3389/fnbot.2018.00043
Ting LH (2007) Dimensional reduction in sensorimotor systems: a framework for understanding muscle coordination of posture. Prog Brain Res 165:299–321
Latash ML (2008) Synergy. Oxford University Press, New York
Alnajjar F, Itkonen M, Berenz V, Tournier M, Nagai C, Shimoda S (2015) Sensory synergy as environmental input integration. Front Neurosci 8:436
d’Avella A, Saltiel P, Bizzi E (2003) Combinations of muscle synergies in the construction of a natural motor behavior. Nat Neurosci 6(3):300
Shibata Alnajjar F, Wojtara T, Shimoda S et al (2013) Muscle synergy space: learning model to create an optimal muscle synergy. Front Comput Neurosci 7:136
Gonzalez-Vargas J, Sartori M, Dosen S, Torricelli D, Pons JL, Farina D (2015) A predictive model of muscle excitations based on muscle modularity for a large repertoire of human locomotion conditions. Front Comput Neurosci 9:114
Takei T, Confais J, Tomatsu S, Oya T, Seki K (2017) Neural basis for hand muscle synergies in the primate spinal cord. Proc Natl Acad Sci 114(32):8643–8648
Costa A, Itkonen M, Yamasaki H, Alnajjar F, Shimoda S (2018) A novel approach to the segmentation of semg data based on the activation and deactivation of muscle synergies during movement. IEEE Robot Autom Lett 3(3):1972–1977
Okajima S, Alnajjar FS, Yamasaki H, Itkonen M, García ÁC, Hasegawa Y, Shimoda S (2018) Grasp-training robot to activate neural control loop for reflex and experimental verification. In: 2018 IEEE International conference on robotics and automation (ICRA), pp 1849–1854
Acknowledgements
We are grateful to Toyota Motor Corporation for providing the DsRS device for the experiments.
Funding
We gratefully acknowledge funding by Toyota Motor Corporation. The funding source was not involved in study design; in the collection, analysis or interpretation of data; in the writing of the report; nor in the decision to submit the article for publication.
Author information
Authors and Affiliations
Contributions
MI and SS conceived of the present research. MI, AC, HY, SO, and SS designed and executed the experiments. MI and SS developed the underlying theory. MI performed the computations. MI and SS wrote and edited the manuscript. All authors discussed the results and contributed to the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Itkonen, M., Costa, Á., Yamasaki, H. et al. Influence of bimanual exercise on muscle activation in post-stroke patients. Robomech J 6, 14 (2019). https://doi.org/10.1186/s40648-019-0142-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40648-019-0142-1